 |
|
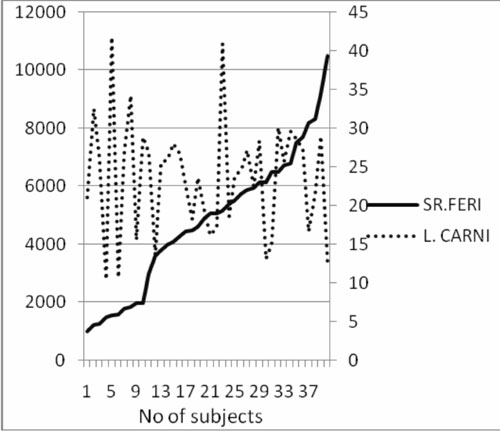
Fig.1
Comparison of L-carnitine
(µM) and ferritin (ng) levels.
|
Discussion
Primary carnitine deficiency is rare although secondary
deficiency may occur due to either decreased biosynthesis, increased
catabolism as in chronic diseases such as thalassemia(7,8), or non
physiologic losses as seen in dialysis(9). In the present study,
thalassemics had significantly lower mean serum total L-carnitine
concentration as compared to controls. Although the precise mechanism of
carnitine deficiency in
b
thalassemia is unknown, it is postulated that this may be related to
reduced hepatic synthesis of carnitine in the liver(7).
In thalassemia, excess iron is deposited in the liver
due to increased absorption and repeated blood transfusion, which could be
responsible for decreased carnitine levels. This iron activates the
respiratory burst and generates superoxide; peroxide and free hydroxyl
radicals(10). L-carnitine is a free radical scavenger which prevents lipid
peroxidation of cell membrane and apoptosis reducing serious tissue
damage(8). Furthermore, it induces HbF synthesis by increased gamma chain
production and decreased alpha chain precipitation, which is the
triggering mechanism for apoptosis. This decrease in apoptosis on
carnitine supplementation was more pronounced in well chelated patients as
compared to those with improper chelation(6). Serum ferritin determines
the chelation status of the patients and is an indirect method of
measuring excess iron deposited in liver(11). Our observation also
suggests that increased liver iron may be responsible for decreased
carntitine levels.
In healthy individuals, carnitine homeostasis is
maintained through endogenous biosynthesis, absorption from dietary
sources, and elimination and reabsorption by the kidney. Nutritional
carnitine deficiency has not been identified in healthy individuals
without metabolic disorders, suggesting that most people can synthesize
enough carnitine(12); even strict vegetarians (vegans) show no signs of
carnitine deficiency despite the fact that most dietary carnitine is
derived from animal sources (13). In this study, though the levels of
carnitine in non vegetarians were higher than in the vegetarians, the
difference was statistically not significant, suggesting that diet alone
is not responsible for alteration in carnitine homeostasis.
As L-carnitine is an essential element for energy
metabolism in mitochondrial fatty acid oxidation, it is most concentrated
in tissues that use fatty acids as their fuel, such as skeletal and
cardiac muscles. Supplementation with L-carnitine has shown not only to
significantly improve cardiac function (14,15) but also general well being
and exercise tolerance(6,7). In our study, no significant difference was
noted between the mean carnitine levels of those who have any clinical
evidence of myalgia, chronic fatigue or muscle weakness as compared to
mean levels of those who were asymptomatic.
Studies are needed to document the role, if any, of
carnitine supplementation in thalassemics.
Funding : None.
Competing interest : None stated.
|
What This Study Adds?
• Carnitine levels are
lower in thalassemics with ferritin>2000ng/dL and in vegetarians.
|
References
1. Shiner E, Rachmilewitz EA. Oxidative denaturation of
red blood cells in thalassemia. Semin Hematol 1990; 27: 70-76.
2. Stamatoyannopoulos JA, Nienhuis AW. Therapeutic
approaches to hemoglobin switching in treatment of hemoglobinopathies.
Annual Rev Med 1992; 43: 497-521.
3. Perrine SP, Miller BA, Faller DV, Cohen RA,
Vichinsky EP, Hurst D, et al. Sodium butyrate enhances fetal globin
expression in erythroid progenitors of patients with HbSS and beta
thalassemia. Blood 1989; 74: 454-459.
4. Walter H. L-Carnitine. Arch Dis Child 1996; 74:
475-478.
5. Palmieri L, Ronca F, Malengo S, Bertelli A.
Protection of b thalassemic
erythrocytes from oxidative stress by propionyl carnitine. Int J Tissue
React 1994; 16: 121-129.
6. El-beshiawy A, Seoud H, Ibharim A, Youssary I, Gabre
H, Ismaeel H, et al. Apoptosis in thalassemia major reduced by a
butyrate derivative. Acta Haematol 2005; 114: 155-159.
7. Tsagris V, Laipi-adamidou G. Serum carnitine levels
in patients with homozygous beta thalassemia: a possible new role of
carnitine? Eur J Pediatr 2005; 164: 131-134.
8. Toptas B, Baykal A, Yesilipek A, Isbir M, Kupesiz A,
Yelcin O, et al. L-carnitine deficiency and red blood cell
mechanical impairment in b thalassemia
major. Clin Hemor Microcircu 2006; 35: 349-357.
9. Evangeliou A, Vassopoulous D. Carnitine metabolism
and deficit- when supplementation is necessary? Curr Pharm Biotechnol
2003; 4: 211-219.
10. Gutteridge JM, Hallwell B. Iron toxicity and oxygen
radicals. Clin Hematol 1989; 2: 195-256.
11. Maris TG. Myocardial and liver iron status
using a fast T*2 quantitative MRI (T*2qMRI) technique. Magn Reson Med
2007; 57: 742-757.
12. Rebouche CJ. Carnitine. In: Shils ME, Shike
M, Ross AC, Caballero B, Cousins RJ. Modern Nutrition in Health and
Disease. 10th ed. Philadelphia: Lippincott, Williams & Wilkins;
2006.p.537-544.
13. Lombard KA, Olson AL, Nelson SE, Rebouche CJ.
Carnitine status of lactoovovegetarians and strict vegetarian adults and
children. Am J Clin Nutr 1989; 50; 301-306.
14. Ei-Beshlawy A, Ragab L, Fattah A, Ibrahim I, Hamdy
M, Makhlouf A, et al. Improvement of cardiac function in
thalassemia major treated with L- Carnitine. Acta Haematol 2004;
111:143-148.
15. El-Beshlawy A, El- Accaoui R, El- Sattar M, El-Deen
M, Youssary I, Shaheen N, et al. Effect of L- Carnitine on physical
fitness of thalassemic patients. Ann Hematol 2007; 86: 31-34.